alpham = -3.48(Vm-11.0)/{exp[(Vm-11.0)/-12.94]-1}b. Canales axónicos:betam = 0.12(Vm-5.9)/exp[(Vm-5.9)/4.47]-1}
alphah = 3/exp[(Vm+80)/10]
betah = 12/{exp[(Vm-77)/-27]+1}
Canal Ca++ (Ca_W):
ICa = gCas2r(Vm-ECa)
ECa = -13.3Ln([Ca,1]in/1200)
alphas = -0.16(Vm+26.0)/{exp[(Vm+26)/-4.5]-1}
betas = 0.04(Vm+12)/exp[(Vm+12)/10]-1}
alphar= 2/exp[(Vm+94)/10]
betar = 8/{exp[(Vm-68)/-27]+1}
Canal de K+ tipo C (K_C_Wtn):
IC = gCc2d(Vm-EK)
EK = -85 mV
alphac = -0.0077(Vm+Vshift+103)/{exp[(Vm+ Vshift +103)/-12]-1}
betac = 1.7/exp[(Vm+Vshift+237)/30]
Vshift = 40Log([Ca,1]in)-105
tauc = 1.1 ms
alphad = 1/exp[(Vm+79)/10]
betad = 4/{exp[(Vm-82)/-27]+1}
Canal de K+ tipo AHP (K_AHP_Wtn):
IAHP = gAHPq(Vm-EK)
EK = -85 mV
alphaq = 0.0048/exp[(10Log([Ca,2]in-35)/-2]
betaq = 0.012/exp[(10Log([Ca,2]in+100)/5]
tauq = 48 ms
Canal de K+ tipo M (K_K_W):
IM =gMu2(Vm-EK)
EK = -85 mV
alphau = 0.016/exp[(Vm+52.7)/-23]
betau = 0.016/exp[(Vm+52.7)/18.8]
Canal de K+ tipo A, proximal (K_A_3):
IAp =gApa4b(Vm-EK)
EK = -85 mV
ainf = 1/{exp[(-Vm-5)/10]+1}
taua = 0.15ms
binf = 1/{exp[(Vm+56)/8]+1}
taub = 5ms if Vm<-30 mV and 5+0.26(Vm+30) if Vm>30 mV
Canal de K+ tipo A, distal (K_A_4):
IAd =gAda4b(Vm-EK)
EK = -85 mV
ainf = 1/{exp[(-Vm-15)/8]+1}
taua = 0.15ms
binf = 1/{exp[(Vm+56)/8]+1}
taub = 5ms if Vm<-30 mV and 5+0.26(Vm+30) if Vm>30 mV
Canal de K+ tipo DR (K_DR_W):
IDR =gDRn4(Vm-EK)
EK = -85mV
alphan = (-0.018Vm)/[exp(Vm/-25)-1]
betan = 0.0036(Vm-10)/{exp[(Vm-10)/12]-1}
Canal de K+ tipo DR (K_DRA_T):c. Dinámica del calcio:IDRA =gDRAnax4(Vm-EK)
EK = -85mV
alphan(ax)= -0.03(Vm+47.8)/{exp[(Vm+47.8)/-5]-1}
betan(ax) = 0.45exp[(Vm+53.0)/-40]
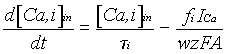
donde taui es la constante de tiempo del "pool" i, fi la fracción del flujo de Ca+ que afecta a cada "pool" (f1=0.7, f2=0.024), w el espesor de la capa de difusión (1 µm), A el área del compartimento, z la valencia del calcio (2) y F is la constante de Faraday.